Génétique et vie en société chez les Hyménoptères (Apis mellifera)
Cet article présente les modèles expliquant l'importance de la génétique des abeilles sur leur vie en société. Il traite de la ploïdie des ouvrières, de la reine et des faux-bourdons ainsi que du rôle de la ploïdie et de l'hétérozygotie dans la détermination du sexe des abeilles.
Le taxon des Hyménoptères compte plus de 120 000 espèces. Il est caractérisé par l'existence de mâles haploïdes (possédant des cellules avec un seul exemplaire de chaque chromosome) et des femelles diploïdes (possédant des cellules avec deux exemplaires de chaque chromosome). La ploïdie correspond au nombre d'exemplaires de chromosomes contenus dans une cellule.
Les Hyménoptères regroupent des espèces eusociales comme dans la famille des Apidea (abeilles) et des Formicidea (fourmis). On retrouve cependant des espèces solitaires chez les Hyménoptères (exemple : Andrènes...).
Les eusociétés sont définies par la superposition, dans une même société :
- de plusieurs générations d'adultes ;
- d'une forte cohésion des membres (échange d'informations et de matière entre les membres) ;
- d'une division des rôles avec spécialisation des membres en "castes" ;
- d'un élevage coopératif de la progéniture.
Cet article traitera de l'influence de la génétique sur la vie en société des Hyménoptères et plus particulièrement du cas d'Apis mellifera (abeille à miel).
Dans la première partie, le calcul du coefficient de parenté entre les différents membres d'une eusociété (colonie) sera présenté. Il sera utilisé, dans la seconde partie, afin d'expliquer la présence d'une reine dans les colonies (seul individu femelle ayant une descendance). Enfin, la relation entre la génétique des abeilles et l'existence mâles sera traitée dans la troisième partie.
Coefficient de parenté et type de reproduction
Chez Apis mellifera, la reine se fait féconder au début de sa vie (entre le 8ème et le 10ème jour après l'éclosion) par environ 15 mâles (polyandrie).
Le coefficient de parenté entre deux individus (1 et 2) représente la probabilité de trouver un gène d'un des deux individus chez l'autre. La valeur du coefficient de parenté varie, suivant que l'on cherche la probabilité de trouver un gène de l'individu 1 chez l'individu 2 ou de l'individu 2 vers l'individu 1, dans le cas où les individus 1 et 2 ne sont pas tous les deux hétérozygotes.
Pour des raisons didactiques, le coefficient de parenté sera d'abord présenté dans le cas d'une fécondation par un seul mâle (monoandrie).
Monoandrie
La monoandrie est un mode de reproduction sexuée où un individu femelle s'accouple avec un seul mâle. Le schéma ci-dessous décrit le calcul du coefficient de parenté chez Apis mellifera. Les rectangles symbolisent les chromosomes de chaque individu, les cases symbolisent les gènes portés par chaque chromosome (portion de la molécule d'ADN contrôlant un caractère particulier). Les cases présentent différentes couleurs qui symbolisent les allèles (versions d'un gène).
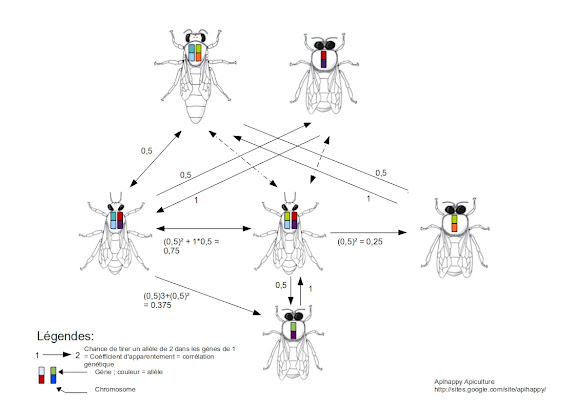
Calcul du coefficient de parenté entre une fille ouvrière (génération II) et sa mère (génération I)
La probabilité de tirer un allèle de la mère chez sa fille est de 0,5 car la fille ouvrière, issue de la fécondation, possède la moitié des gènes de sa mère et la moitié des gènes de son père. Il y a donc 1 chance sur 2 (½ =0,5) de tirer un allèle de la mère chez la fille ; le coefficient de parenté mère-fille est alors de 0,5.
Calcul du coeficient de parenté entre deux soeurs ouvrières (génération II)
Il faut chercher la probabilité de tirer le même allèle provenant de la mère (a) plus la probabilité de tirer le même allèle provenant du père (b) d'une soeur chez l'autre soeur.
a) La probabilité que les deux soeurs possèdent le même allèle en provenance de leur mère est égale :
- à la probabilité de tirer un allèle de la mère chez sa fille (0,5 car elle possède la moitié des gènes de sa mère et la moitié des gènes de son père) ;
- que l'on multiplie à la probabilité de tirer le même allèle chez l'autre soeur (0,5 car les femelles sont diploïdes, elles possèdent deux exemplaires d'un même chromosome et donc deux exemplaires d'un même gène ; ce gène peut potentiellement être présent sous deux versions : allèles). Il y a donc 1 chance sur 2 (½ =0,5) de tirer un allèle d'une soeur chez l'autre soeur.
La probabilité que les deux soeurs possèdent le même allèle en provenance de leur mère est donc égale à 0,5 x 0,5 = 0,5².
b) Il faut additionner à ce résultat la probabilité que les deux soeurs aient le même allèle en provenance du père soit :
- la probabilité de trouver un allèle du père chez sa fille (0,5 car elle possède la moitié des gènes de sa mère et la moitié des gènes de son père)
- que l'on multiplie à la probabilité de trouver ce même allèle chez l'autre soeur (1 car les mâles sont haploïdes, ils ne possèdent qu'un seul exemplaire de chaque chromosome et donc qu'un allèle pour chaque gène ; un allèle transmis à une fille sera le même pour toutes les filles).
La probabilité que les deux soeurs aient le même allèle en provenance du père est donc égale à 0,5 x 1 = 0,5.
Le coefficient de parenté entre deux soeurs est donc de 0,75.
Polyandrie
Généralement les reines sont fécondées par plusieurs mâles : ces modalités de fécondation influencent la valeur du coefficient de parenté entre les individus d'une même colonie.
Prenons l'exemple d'une fécondation par deux mâles. Dans ce cas, il y a deux fois moins de chances de tirer le même allèle chez deux soeurs ouvrières. Pour obtenir le coefficient de parenté entre deux soeurs ouvrières en fonction du nombre de mâles fécondants (génétiquement différents), il suffit de diviser la chance de tirer un allèle du père chez la fille (0,5) par le nombre (K) de mâles fécondants (génétiquement différents). La formule est donc : coefficient de parenté entre deux ouvrières issues de polyandrie = (0,5)² + (0,5/K)

Le graphique suivant, obtenu à partir de l'équation précédente, présente l'évolution du coefficient de parenté en fonction du nombre de mâles fécondants génétiquement différents. Il ne faut pas oublier que dans l'ensemble des mâles fécondants, certains peuvent être génétiquement identiques.
La probabilité de tirer un allèle de la soeur chez son frère est toujours égale à (0,5)2 car les mâles n'ont pas de père. Le coefficient de parenté soeur-frère est donc égal :
- à la probabilité de tirer un allèle de la mère chez sa fille (0,5) ;
- que l'on multiplie à la probabilité de tirer le même allèle chez le frère (0,5 car les mâles possèdent un des deux allèles de la mère pour chaque gène et la soeur la moitié des allèles de son père et la moitié des allèles de sa mère).
La probabilité de tirer un allèle de la soeur chez son frère est de 0,5 x 0,5 = (0,5)2.
Ce graphique met en évidence que le coefficient de parenté soeur-soeur est toujours plus important que le coefficient de parenté soeur-frère. Cependant, lorsque le nombre d'accouplements augmente, le coefficient de parenté entre deux ouvrières de la ruche diminue puisque celles-ci ont la même mère mais ne sont pas forcément issues du même mâle fécondant.
Bilan
- Les mâles n'ont pas le même père mais seulement un grand père car ils sont (majoritairement) issus d'un oeuf non fécondé de reine.
- Les ouvrières d'une colonie sont regroupées en fratries (ensemble d'individus étant issus d'un même père). Deux ouvrières de deux fratries différentes sont donc des demi-soeurs (s.s.).
Génétique et présence d'une reine
Pondre ou l'altruisme fraternel (sélection de parentèle)

En 1964, le biologiste William Donald Hamilton émet la théorie de la sélection de parentèle. Elle explique l'origine des comportements altruistes au sein des eusociétés d'abeilles.
Des individus frères ou soeurs possèdent en partie les mêmes allèles, provenant de leur mère ou de leur père. Pour un individu appartenant à une eusociété, aider ses frères et sœurs revient donc "à maintenir, de manière indirecte, une partie de ses allèles dans la colonie".
Une ouvrière a plus de gènes en commun avec sa sœur (0,75) qu'avec sa descendance (0,5). Dans ce cas, le comportement d'altruisme fraternel est favorisé, ainsi que les allèles contrôlant ce comportement. Sans être finaliste, on peut simplifier en disant qu'il est alors plus avantageux d'aider sa sœur que de se reproduire pour transmettre ses allèles.
Dans le cas d'une reproduction multiple, il devient plus avantageux pour une ouvrière de se reproduire au-delà de deux accouplements de la reine. Pour palier à se problème, les oeufs des ouvrières sont éliminés.
L'élimination des oeufs d'ouvrière
Une ouvrière est très éloignée génétiquement de la descendance de ses sœurs (37,5 %). Cette différence est particulièrement accentuée lorsque le nombre de mâles fécondants est important. Cette observation semble être la clé expliquant le comportement de destruction du couvain des ouvrières. De plus, il n'est pas "rentable" de pondre des oeufs pour une ouvrière car ils seront détuits par ses soeurs.
Génétique et proportion des mâles
Polyandrie et proportion des mâles
Ce modèle permet d'expliquer les variations de l'abondance des mâles dans une colonie. Plus l'abondance de mâles fécondants est importante, plus la différence entre le coefficient de parenté sœur/sœur et sœur/mâle diminue. Il devrait donc avoir un fort taux de mâles dans les colonies lors de fécondations multiples.
Cette conclusion ne semble pas correspondre aux observations de terrain. Il existe peut-être d'autres mécanismes contrôlant l'abondance des mâles.
Consanguinité et proportion des mâles
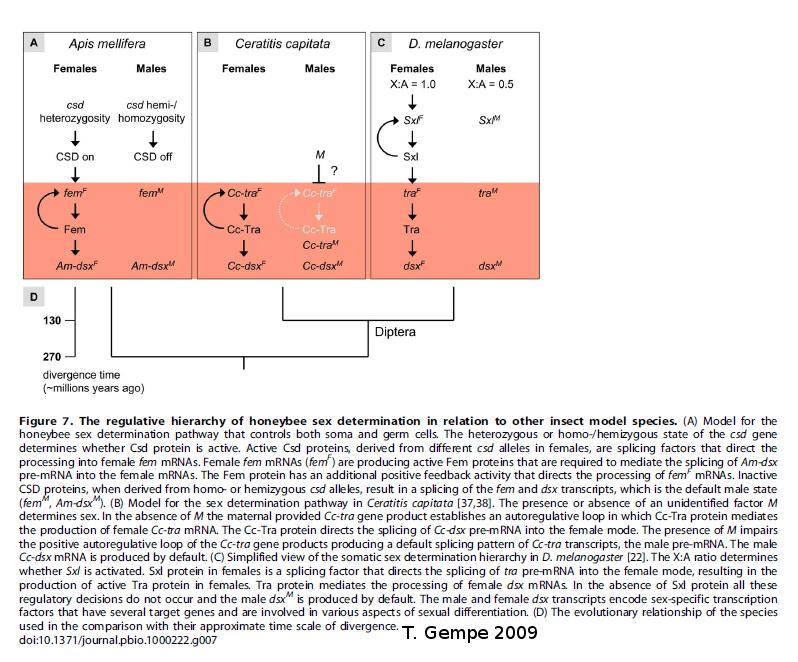
Le document ci-dessous présente les résultats des travaux de Gempe sur les mécanismes de détermination du sexe chez les Hyménoptères. Cette étude révèle qu'il existe des mâles diploïdes.
La détermination du sexe des abeilles est contrôlée par l'homozygotie du gène csd :
- les abeilles homozygotes, dont les allèles sont identiques sur le gène csd, sont des mâles ;
- les abeilles hétérozygotes, dont les allèles du gène csd sont différents, sont des femelles.
Les abeilles haploïdes ne peuvent pas être hétérozygotes : il s'agit donc de mâles.
Quinze allèles sexuels existent au minimum. Cette diversité permet de limiter les cas d'homozygotie sur le gène csd. Cependant, ce mécanisme de détermination du sexe chez les abeilles explique la consanguinité et le cas des ruches bourdonneuses. La consanguinité augmente la proportion d'homozygotes dans la population.
Conclusion
Le modèle théorique, expliquant le maintien du comportement fraternel des abeilles au cours de l'évolution, permet de comprendre plusieurs comportements. Il ne faut cependant pas oublier qu'un modèle est une simplification de la réalité : c'est donc un outil de compréhension.
Pour aller plus loin:
Comment analyser le pollen de mon miel?
Comment estimer le nombre d'abeilles sur un cadre?
Connaitre le vocabulaire d'apiculture







Essaim d'abeilles Buckfast Dadant Essonne Yvelines
- Promo !
- -18,01 €